| Редакция
| Авторы | Форум
| Гостевая книга |
Текущий номер |
Яков Гельфандбейн
<ИЛИ-ИЛИ> ИЛИ <И-И>, ИЛИ О ШАРЛАТАНСТВЕ В ОНКОЛОГИИ.
<Паразитарная> гипотеза возникновения и развития рака с момента опубликования
работ Т.Свищевой заняла весьма агрессивную позицию, противопоставляя себя
т.н. <генетической> теории и методам лечения официальной медицины.
Ее сторонники, не имея доказательной аргументации, ссылаясь на сомнительные
факты и цифры, почерпнутые из cомнительных источников, а также на сомнительный
опыт столь же сомнительных <народных целителей>, не стесняясь в выражениях,
стремятся доказать <зловредность> врачей - онкологов, генетики в целом
и отсутствие научно обоснованной методологии борьбы со злокачественными
опухолями.
Поток дешевых публикаций захлестнул книжные рынки и СМИ, неряшливая статистика и крикливость <доказательства> исключительности паразитарного происхождения рака, а заодно инфаркта, инсульта, тромбоза и прочих сердечных заболеваний, и взбудоражили общественное мнение.
Опасность заключается в том, что такие публикации
могут убедить некомпетентного читателя в истинности этих "чудесных
открытий".
Предлагая очередное "снадобье", авторы пишут о том, что нельзя
применять химиотерапию и лучевое лечение, возможно, обрекая тем самым больного
на трагический конец! (см. Н.Н. Трапезников, академик РАН и РАМН, Л.А. Дурнов,
академик РАМН, <Шарлатанство и онкология> (по поводу очередного "великого
открытия").
Основой паразитарной гипотезы является утверждение, что злокачественная
опухоль образуется не как специфическое состояние клеток биологического
субстрата, а представляет собой незаживающую рану, образованную жизнедеятельностью
колонии паразитов - трихомонад. Из этого делаются весьма сомнительные выводы
о методах лечения и рекомендуются такие средства, как "обтирание тела
спермой" или "калом лошади, взятым на лету", регулярное питье
мочи, употребление керосина с применением различных примесей, что само по
себе абсурдно. В прессе публиковались сообщения о лечении рака с помощью
водки, масла и лампы Чижевского. Издана даже книга о лечении рака пивом.
Опасность заключается в том, что такие публикации могут убедить некомпетентного
читателя в истинности этих "чудесных открытий". Так как речь идет
о здоровье людей, такие книги и публикации просто преступны.
Различить трихомонаду можно только с помощью специальных
микроскопов. Опытный исследователь, даже если трихомонада "не бросается
в глаза" в первый момент, "чувствует" её по специфическому
"грязному" фону, "рваным" эпителиальным клеткам с околоядерным
просветлением, кокковой флоре, большому количеству лейкоцитов и... чему-то
необъяснимому, но интуитивно ощущаемому. Среди множества ее особенностей
- неразличимость от эпителиальной клетки, способность не просто повторять
ее рельеф, но и проникать в клетку хозяина.
Кроме того, она способна защититься от разрушительного действия организма
и прикрепляться к лейкоцитам, эритроцитам, эпителиальным клеткам за счет
сложного белка, содержащего углеводные компоненты, - гликопротеида.
Неразличимость трихомонады на предметном стекле обычного
микроскопа от эпителиальной клетки, создает основу для спекуляций. Отсутствие
количественной определенности паразитарного поражения, трактуемого как опухоль,
недостаточно уверенного понимания имеющихся фактов и искаженной их трактовки,
делает необходимым выработку научно обоснованного базиса, позволяющего осмыслить
механизмы такого поражения. Это связано с претензиями паразитарной гипотезы
на исключительность, отрицающую самую альтернативность, как выбор понимания
этого опаснейшего поражения человека и необходимостью блокирования применения
сомнительных средств борьбы с ним.
Выработка такого базиса необходима и для перевода разгоревшейся полемики
от спора интересов в русло биологических и смежных наук.
Здесь мы прокомментируем некоторые результаты математического исследования, направленного на оценку правдоподобности самой гипотезы паразитарного происхождения рака в условиях фактического незнания провозглашенных паразитарных механизмов. Мы будем ссылаться на хорошо определенную математическую модель <клеточного> рака, как на эталон для сопоставления.
Постараемся ответить на главный вопрос: есть ли рак
состояние озлокачествленных клеток или это продукт жизнедеятельности паразитов,
не имеющий никакого отношения к клеткам? С другой стороны, не существуют
ли оба процесса одновременно, обуславливая друг друга? Это означает, что
мы рассматриваем двойную дилемму - отражает ли утверждение о паразитарности
рака отношение альтернативности <ИЛИ(клетка) - ИЛИ(паразит)>, или
отношение совместности <И(клетка) - И(паразит)>? От ответа на этот
вопрос во многом зависит тактика и стратегия борьбы с грозным заболеванием.
Так как ролевая функция трихомонады гипотезой не конкретизируется, то такое умолчание дает основания предполагать ее двойственную роль - и как агента влияния, и как агента реализации процесса малигнизации. Это умолчание очень похоже на скрытую подмену понятием <происхождение> понятия <исполнения>, допускающую манипуляцию терминами.
Из этого следует необходимость оценки роли паразита
как агента влияния, определяющего генезис (происхождение) процесса, и одновременно
(основополагающего в обсуждаемой гипотезе) утверждения, что именно паразитарное
<происхождение> образует исключительную форму рака - вообще не связанную
с клеточными преобразованиями. Отмеченная подмена понятий требует оценки
и его вероятной роли в качестве <реализующего>, исполняющего
механизма, своеобразного динамического <промотора> - активатора процесса
именно на клеточных множествах. Не исключено также, что паразит создает
среду некоторого <усилителя> устойчивости, той части механизма озлокачествления,
который делает процесс малигнизации необратимым.
Заметим, что утверждение о вероятности паразитарного
происхождения процесса инициализации не отвергается и результатами косвенных
математических исследований клинических материалов по человеку и лабораторных
исследований
на животных, о чем мы неоднократно писали еще в 70-х годах. Более того,
оно вытекает из них. Аргументы авторов гипотезы не дают также оснований
категорически отрицать паразитарное присутствие в клинике раковых опухолей.
Это создает иллюзию корреляции (вероятностной связи) <присутствия> и усиливает отмеченную выше необходимость исследования возможности непосредственного включения в осуществление процесса опухолеобразования специфических паразитарных механизмов.
К сожалению, будучи сторонником количественной определенности, мы вынуждены ограничиться логическими рассуждениями, из-за полного отсутствия информации не только по паразитарным воздействиям, но и по морфологической картине их реализации. Попытки войти в контакт с апологетами паразитарной теории и получить нужную информацию оказались безуспешными. Это создает лишний повод сомневаться в правдивости их громких заявлений, утверждающих цели борьбы в защиту <истины>.
Пути и направления такого рода исследований разнообразны Самые достоверные выводы могут быть получены в результате специально поставленных экспериментов. В то же время использование имеющихся результатов моделирования способно, образуя метод логического анализа, доставить знания, недоступные для других методов, одновременно определяя пути дальнейших исследований, если выявленная (хотя бы на качественном уровне) корреляция присутствия, покажется существенной и действительно образует корреляцию влияния. В первую очередь это касается возможностей выявления в условиях паразитарного влияния механизмов естественно-технологических процессов управления на клеточном уровне и его отношений с процессами белкового синтеза на молекулярном уровне, т.е. межуровневых отношений.
Мы опирались на следующее исходное гипотетическое
рассуждение (его понимание - ключ к пониманию всех последующих умозаключений):
1)в том случае, если злокачественное поражение клеток не коррелируется с
паразитарным раком, модель межуровневых отношений при наличии опухоли не
отразит корреляции статистики клеточных множеств и динамики белкового
синтеза. Это соответствует предположению о роли паразита исключительно как
агента, возбуждающего процесс, но не участвующего в нем;
2)если модель межуровневых отношений в условиях паразитарного поражениявсе-таки
отразит эти отношения, то это будет означать, что паразитарный рак опосредуется
через эти отношения и что роль паразита как агента влияния двойственная:
как возбудителя и как агента влияния, осуществляющего протекционную функцию
активатора активности - динамический <промотор>.
При этом характер протекционизма и параметры его проявления, формируются в зависимости от характера агента влияния - конкретного паразита.
Для решения задачи мы предположили, что роль реципиента, принимающего воздействия возбуждения, отводится участку в геноме, на котором входные воздействия в виде сигнальных молекул (токсинов жизнедеятельности паразитов) активируют первый этап реализации генетической информации в клетке, в процессе которого последовательность нуклеотидов ДНК <переписывается> в нуклеотидную последовательность РНК. В этом случае молекулы токсинов играют роль сигнальных молекул белков, и это может отражать либо их заместительные, либо родственно-ролевые отношения. Роль адресата исполнителя воздействия возбуждения, отводится механизмам последующих этапов биосинтеза белков.
Полученные нами результаты в трактовках понятий математики,
естественно, могут быть трудны для неподготовленного читателя. Поэтому в
нашем комментарии мы нашли возможным обойти технологические подробности
сопоставительных процедур на моделях и только дать краткие разъяснения полученных
результатов при самом минимальном применении специальных терминов.
Автор заранее приносит свои извинения у тех, кто может определить такое
изложение, как упрощенчество.
Мы имеем хорошо обусловленную и наглядную модель,построенную по экспериментально-клиническим данным, на которой отражены все особенности процесса опухолеобразования. Такая модель играет роль эталона, на котором можно сопоставлять самые различные гипотезы о генезисе рака. Ее отличительной особенностью является количественная определенность всех изменений, происходящих на множествах клеток в любой момент времени, которые мы не можем видеть и оценить даже "вооруженным" глазом. Более того, модель отражает отклонения от нормы морфологии клеточного поля, как результат отклонения управлений на клеточных множествах, связанных с изменениями отношений на молекулярном уровне.
Это означает, что модель одновременно является и
средством количественного отображения системы межуровневых молекулярно-клеточных
отношений порождения между (ненаблюдаемыми) порождающими процессами синтеза
белков на молекулярном уровне, и порожденными (наблюдаемыми) изменениями
феноменологии клеточного поля. Внося количественную определенность, модель
отражает основной закон развития - накопления малых количественных изменений
и их перехода в скачкообразные качественные. Такие качественные изменения
отражаются, в частности, некоторым набором цифровых значений специфических
признаков - <констант состояния>, являющихся его идентификаторами.
Константы состояния - визитная карточка опухоли. В процессе опухолеобразования,
сформировавшись, константы остаются неизменными, играя роль <инвариантов>.
Модель представляет процесс опухолеобразования в
виде объекта, обладающего входом и выходом, реализующего все множество механизмов
этого естественно-технологического процесса. Выход отображает наблюдаемый
феномен изменений клеточного поля, вход отражает воздействие носителя влияния(паразита),
возбуждающего процесс сигналом, поступающим на <реципиент> - приемник
воздействия. Механизмы процесса описываются множеством математических выражений,
отражающих те или иные преобразования ненаблюдаемого входа, которые приводят
к наблюдаемому выходу. Множества таких математических выражений преобразуются
в геометрическое отображение.
Нас интересует выявление с помощью такой модели характера входа - того агента
влияния, который, по предположению, запускает, а быть может, и протекционирует
процесс, и это возможно, трихомонада или иной паразит.
Т.Свищева называет 30 видов микроорганизмов, 15 видов
паразитов, 96 видов вирусов, 40 видов грибков, участвующих, по ее мнению,
в процессе опухолеобразования. Особое место, по мнению Елисеевой, занимают
среди паразитов трихомонада (<мать рака>), а среди грибков - микозис
фунгоидес <отец рака>). Таким образом, рак постулируется как незаконное
дитя паразита и грибка. Но незаконные, как и законные дитяти, - бывают разные,
и способ их жизнедеятельности различен, да и не понятно, вместе ли "родители"
"работают" или они "в разводе". Елисеева встречала их
(не указывая - совместно или врозь) в 100% случаев злокачественного поражения,
подчеркивая, что именно с появлением <отца> опухоль становится злокачественной,
а, как отмечает В.Плюснин, <отходами этой семейки питается прочая шушера>.
Поэтому нам далеко
небезразлично, каким образом осуществляется паразитарный запуск процесса
и, что важно, является ли функция запуска единственной функцией паразита
или же паразит, запустив процесс, продолжает его протекционировать и при
этом действуют ли паразиты поврозь или совместно, с помощью или без помощи
своих сотоварищей по разбою.
В условиях полного отсутствия количественно определенной информации нет доказательных оснований полагать, что естественно-технологические механизмы реализации относительно агента влияния существенно различимы по своему характеру. На начальном этапе анализа возможны как предположение об единстве их представления, так и просто оговорка о рассмотрении гипотетически возможного варианта.
Мы предположили, что гипотеза паразитарного рака
справедлива, ибо без такого допущения попросту исчезает предмет исследования.
Кроме того, мы гипотетически посчитали, что агент влияния в общем случае
образуется именно паразитом, способным не только запускать процесс инициации
рака, но и создавать среду его поддержания и развития, т.е решали задачу
в общем виде.
Эту вторую его способность мы определили как функцию <протекционизма>,
реализующего цели динамического промотора. При этом, следуя паразитарной
гипотезе, мы учли, что колонии паразитов дистанцируются от тела опухоли,
и обсудили это предположение на вероятности ложной идентификации паразитов
при построении модели, как отражения процесса озлокачествления именно клеток
биологического субстрата. По данным эксперимента было установлено, что отображения
<ложных> клеток (неидентифицированных паразитов) в статистике ничем
не отличаются от отображения истинных клеток состояния озлокачествления.
Это обстоятельство важно, так как оно, в известной степени, обосновывает
исходное гипотетическое рассуждение относительно межуровневых отношений,
высказанное выше.
В соответствии с предположением о многофункциональной роли паразита мы рассмотрели естественно-технологический процесс в преобразованном виде, условно представив его в виде двух последовательных подпроцессов - на молекулярном и клеточном уровнях, связав их множеством количественно определенных межуровневых молекулярно-клеточных отношений порождения.
Вход рассматривался как агент влияния, реализующий поступление сигнала влияния на молекулярный уровень. Базируясь на информацию, не связанную с результатом собственного модельного исследования, мы предположили, что сигнал воздействия от агента влияния адресуется как реципиенту хромосоме, ответственной за возникновение рака (в качестве примера - это 13 хромосома, ответственная за рак молочной железы). Такое предположение позволило построить ряд выводов и высказываний, не только хорошо согласующихся с <генетической> теорией, но и с широко декларируемым опытом паразитарной гипотезы. Это представило возможность сформулировать ряд практических рекомендаций по выработке согласованных мер, определяющих стратегию противораковой борьбы для случая, когда положенные в основу гипотезы предпосылки верны.
Основные выводы сводятся к заключению, что паразитарное поражение не вызывает какой-либо <специфической>, исключительной формы рака, определяя только своеобразные формы его порождения и протекционирования. При этом процесс его возникновения и развития на клеточных множествах, возбужденных сигналом агента влияния, определяясь как <типовой>, не имеет видимых внешних различий и отличий механизмов на клеточном поле и может быть объектом всех стандартных и традиционных видов контроля и репрессий.
Вместе с тем, в ходе подавления, в силу недосягаемости
паразита репрессивными воздействиями, направленными на клетки, токсины как
результаты его жизнедеятельности продолжают поступать на реципиенты влияния
непрерывно и независимо от состояния репрессируемого клеточного поля, даже
при полном подавлении озлокачествляющихся клеток. Это означает, что они
так же непрерывно возбуждают рецидивы инициализации и роста, вовлекая в
процесс как еще не возбужденные, так и уже возбужденные клетки (такой процесс
наглядно отображается на модели). Это, в известной мере, определяет роль
паразитарного влияния как <усилителя устойчивости> - части механизма,
который делает процесс малигнизации необратимым. Из этого следует необходимость
непрерывной противопаразитной борьбы до полного и
контролируемого исчезновения сигнала влияния.
Но в любом случае достижение успеха возможно только до определенного момента развития процесса. Такой момент соответствует переходу от пластического состояния клеточного поля к гиперпластическому состоянию. Этот момент отчетливо отражается на всех видах и вариантах математической модели своей константой состояния как момент определенного взаимодействия специфических клеточных множеств. Необходимо отметить, что все упомянутые константы, одновременно являющиеся инвариантами и идентификаторами состояния, играют большую роль в задаче мониторинга процесса.
Таким образом, исключительное место паразитарного рака в онкологии элиминируется. Его специфичность достаточно очевидно не подтверждается, ибо феноменология процесса отражает клеточные, а не иные изменения.
Декларируемая исключительная способность к излечению - видимая. Она не определяется только фактором устранения его функций влияния и протекционирования: их прерывание никак не прекращает установившегося процесса именно клеточного, а не иного роста. Паразитарный рак, как специфическая форма опухолеобразования, уверенного подтверждения на моделях не находит. Он может квалифицироваться как своеобразная форма инициализации и протекционирования. Двойная дилемма <ИЛИ-ИЛИ> или <И-И> разрешена в пользу отношения совместности <И-И>: клеточный рак может совмещаться (а может и не совмещаться) с опухолеобразованием паразитарного происхождения, более того,паразитарная среда может инициировать клеточный рак. Паразитарное поражение отношения альтернативности не образует.
Здесь мы должны оговориться: сказанное выше соответствует имеющейся в нашем распоряжении информации. По мере накопления верифицированных количественно определенных исходных данных по множествам элементов, образующих тело опухоли в условиях паразитарного поражения, некоторые из приведенных точек зрения, даже для принятых гипотетических предпосылок, могут измениться.
Неудачи излечения стандартными методами, наблюдаемые на стадиях инициализации и развития процесса при паразитарном поражении скорее всего связаны с пренебрежением мониторингом - постоянным слежением за текущим состоянием клеточного поля с ОДНОВРЕМЕННЫМ контролем паразитарного присутствия. Отсутствие мониторинга исключает борьбу с постоянно осуществляемым воздействием агента паразитарного влияния, так как при стандартных репрессиях онкогенов паразит остается вне пределов досягаемости. В то время, когда озлокачествленные клетки так или иначе поражаются, паразитарное влияние сохраняется как дистанционное, приводящее к непрерывному возбуждению рецидивов. При этом не исключено, что в связи с изменением паразитами вида выделяемых ими токсинов или благодаря усилиям <семейки прочей шушеры>, эти рецидивы могут возникать в виде различных форм опухолей. В качестве средств мониторинга могут быть использованы компараторные математические модели с автоматическим вводом модельных данных текущего состояния, основанные на различных его представлениях, с вычислением текущих данных для проведения сопоставительных процедур. Такие модели и технические устройства известны.
С другой стороны, провозглашаемые апологетами паразитарной теории успехи, основанные на уничтожении трихомонадных колоний, действительно могут иметь место при сочетании традиционных репрессивных возможностей и средств подавления паразитарного влияния в условиях, когда развитие процесса еще не вышло за рамки перехода к гиперпластическому состоянию. Выход за пределы этого состояния означает включение собственных клеточных, а быть может, одновременно и <промоторных> средств сохранения устойчивости процесса озлокачествления ("раковый гомеостат"), что делает процесс опухолеобразования необратимым при любых внешних воздействиях, любыми средствами его прерывания, кроме радикальных. Поэтому мониторинг должен обеспечивать и контроль близости к этому состоянию. Программа мониторинга есть продукт математического моделирования.
Интересно, что при этом в ситуации конкурентной борьбы можно создавать иллюзии безусловного подавления рака и этому способствуют неудачи прерывания процессов озлокачествления традиционными методами при отсутствии всестороннего мониторинга и, следовательно, в условиях пренебрежения противопаразитной борьбой. Это связано с тем, что осуществляя мониторинг, мы получаем возможность одновременного контроля и управления состоянием клеточного поля и эффективностью паразитарного влияния на его изменения. Это позволяет репрессировать озлокачествленные клетки традиционными методами при одновременном исключении рецидивов путем параллельного подавления протекционистского, промоторного паразитарного влияния.
При хорошем мониторинге создается видимость преувеличенной эффективности противотрихомонадной борьбы, а традиционные методы подавления процессов роста оставляются "за кадром".
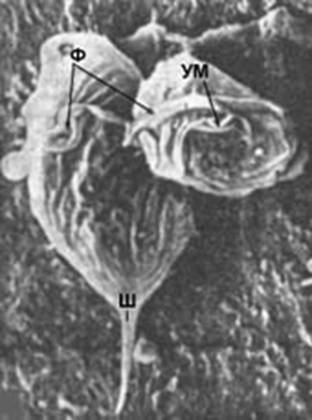
Трихомонада. При электронной микроскопии (ЭМ) видны детали строения (ЭМ х 8000 ).
Ш - «шипик» (жгутик), УМ - ундулируюшая мембрана– волнообразный органоид движения, Ф - фибриллы – нитевидные белковые (опорные) структуры, органоиды (органеллы) движения
Обсудить этот текст можно здесь