| Редакция
| Авторы | Форум
| Гостевая книга |
Текущий номер |
Александр Марков
Переходные формы
И сам Дарвин, и все его последователи прекрасно понимали, что если теория
эволюции верна, и виды действительно превращаются друг в друга, а не сотворяются
богом в раз и навсегда неизменном виде, то в прошлом должно было существовать
множество переходных, промежуточных форм. Фактически, между любыми двумя
видами, сколь бы мало похожими друг на друга они ни были, должен существовать
непрерывный ряд переходов. Однако палеонтология во времена Дарвина не могла
похвастаться большим количеством свидетельств плавного превращения видов
друг в друга. Дарвина очень смущал этот факт, и его объяснению он посвятил
в своем знаменитом труде "Происхождение видов" не один десяток
страниц. Основная его мысль состояла в том, что палеонтологическая летопись
– очень неполная, туда попадает лишь малая часть всех существовавших в прошлые
эпохи видов.
С тех пор палеонтологических находок стало гораздо больше. И во многих
случаях действительно удалось проследить в деталях весь процесс перехода
от одних видов к другим. Особенно в таких группах организмов, которые хорошо
сохраняются в ископаемом состоянии, и к тому же бывают очень многочисленными
– например, у двустворчатых моллюсков с их очень прочными известковыми раковинами.
Оказалось, что, в полном соответствии с гипотезой "прерывистого равновесия",
вид может долго (несколько миллионов лет) существовать в стабильном состоянии,
а потом вступать в короткую фазу "дестабилизации". Отличительная
особенность этой фазы – резкий рост изменчивости. Устойчивая система как
бы расшатывается, внутренние связи слабеют, то есть происходит частичная
дезинтергация – своего рода возврат от порядка к хаосу. Этап "дестабилизации"
не может продолжаться долго, потому что жизнеспособность популяции в этот
период, судя по всему, сильно снижается. Популяции остается либо вымереть,
либо найти какое-то новое устойчивое состояние – в этом случае возникает
новый вид.
Модель "прерывистого равновесия", которую удалось в ряде случаев
подвердить на фактическом материале, хорошо объясняет, почему переходные
формы попадаются палеонтологам реже, чем хотелось бы классическим дарвинистам.
Стабильное существование вида длится гораздо дольше, чем краткий этап "дестабилизации",
во время которого и происходит превращение одного вида в другой, и к которому
приурочены пресловутые "переходные формы". К тому же сам факт
стабильного существования вида показывает, что в это время он находится
в гармонии с окружающей средой. Дестабилизация, напротив, свидетельствует
о кризисе. Поэтому совершенно естественно, что численность вида в стабильные
периоды обычно гораздо выше, чем в период дестабилизации. Что же касается
неполноты летописи, то, несмотря на миллионы новых находок ископаемых организмов,
она по-прежнему весьма велика. По приблизительным оценкам, даже среди тех
организмов, которые сохраняются лучше всех – морских животных с твердым
скелетом – только 10-15% видов, населявших древние моря, стали к настоящему
дню известны палеонтологам. От сухопутных же видов не сохранилось и одного
процента. Ну а мягкотелые животные (за исключением редчайших уникальных
случаев) вообще почти не имеют шансов оставить память о себе в геологической
летописи.
Несмотря на все эти "объективные причины", ситуация с переходными
формами все-таки не совсем понятная. Дело в том, что переходы между отдельными
видами, родами и семействами в пределах одной большой группы обнаружить
удается довольно часто. А вот переходы между самими крупными группами встречаются
гораздо реже. Самая удручающая ситуация – с кембрийским "взрывом",
когда около 570 миллионов лет назад почти одновременно
и словно бы ниоткуда в летописи появилась большая часть современных
типов животных. Это послужило основой появления так называемого "научного
креационизма" (хотя научного в этом учении, честно говоря, мало), который
допускает мелкие самопроизвольные эволюционные преобразования (модификации),
но утверждает, что основные типы и планы строения были все-таки сотворены
богом и не могут видоизменяться под действием мутаций и отбора. Эти "богосотворенные"
типы получили особое название: "барамин"[1].
Во время крупных эволюционных превращений (таких, как появление новых
типов) порой происходят весьма странные вещи, которые трудно заранее вообразить
и смоделировать на основе сравнения известных современных представителей
разных типов животных.
Например, давно считалось общепризнанным, что предками членистоногих (ракообразных,
трилобитов, насекомых, паукообразных, многоножек) были кольчатые черви.
Они похожи на членистоногих по целому ряду признаков. Тело тех и других
состоит из сегментов (члеников) с парными конечностями на каждом
сегменте (у членистоногих это ноги, исходно двуветвистые, как у трилобитов
и многих раков; у кольчатых червей – двуветвистые "параподии"
с хитиновыми щетинками). Нервная система в обоих типах состоит из брюшной
нервной цепочки с утолщениями (ганглиями) в каждом сегменте и кольца, окружающего
глотку (мозг формируется из верхней дуги этого кольца). Сердце располагается
на спинной стороне и гонит кровь вперед. Сходны и ранние этапы индивидуального
развития (спиральное дробление яйца).
Естественно было предположить, что
кольчатые черви "превратились" в членистоногих в результате постепенного
соврешенствования параподий (они постепенно удлинялись, покрывались плотной
хитиновой оболочкой и превращались в членистые ноги) и дифференцировки сегментов
(исходно одинаковые сегменты кольчатых червей группировались и сливались,
образуя голову, грудь, и другие отделы тела членистоногих). Соответственно
и предполагаемые переходные формы в умах ученых приобретали совершенно определенные
очертания. Они – эти воображаемые переходные формы – естественно, наделялись
смесью признаков членистоногих и кольчатых червей (например, все сегменты
одинаковые, но несколько передних уже слились в "голову"; параподии
– еще не членистые, но уже удлиненные и напоминающие ноги ракообразных).
Основные черты строения переходных форм были уже "ясны", оставалось
только найти их ископаемые остатки… И вот тут-то начались сюрпризы.
Поначалу эволюционистов "обнадежила" сприггина – мягкотелое
животное вендского периода (около 600 млн. лет назад), которое вроде бы
имело ожидаемое сочетание признаков: цельную "голову" в форме
полумесяца, напоминающую головной щит трилобита, и длинное тело, состоящее
из одинаковых сегментов с длинными придатками, напоминающее многощетинковых
кольчатых червей.

Сприггина – переходная форма
между кольчатыми червями и членистоногими?
Но если внимательно присмотреться к сохранившимся отпечаткам сприггины,
можно заметить, что конечности у нее располагаются не попарно друг напротив
друга, как у всех без исключения кольчатых червей и членистоногих, а в шахматном
порядке (как если бы левую половину ее тела сдвинули относительно правой
на полсегмента вперед или назад). Такая "билатеральная симметрия со
сдвигом" специалистам по сравнительной анатомии представлялась совершенно
немыслимой. Абсолютно непонятно, как росло тело сприггины, как формировались
сегменты (и вообще – можно ли это назвать сегментами?), как были устроены
ее кровеносная и нервная системы, структура которых определяется механизмом
образования сегментов…
Еще лучше эта "невероятная" сегментация видна у других вендских
форм, напоминающих примитивных членистоногих – например, у вендии.

Вендия похожа на мягкотелого
трилобита, только "ноги" у него расположены не друг напротив друга,
а в шахматном порядке.
Вендия очень похожа на примитивное мягкотелое членистоногое, и все бы
хорошо, если бы не эта невероятная "сдвинутая" симметрия! Надо
сказать, что и все остальные вендские животные, напоминающие кольчатых червей
или примитивных членистоногих, "сдвинуты" таким же образом. Состоят
ли они в родстве с настоящими кольчецами и членистоногими или те произошли
от каких-то других, неизвестных нам предков; возможно ли эволюционное превращение
"сдвинутой" симметрии в нормальную – до сих пор неизвестно.
Что ж, если вендская мягкотелая фауна дала больше новых загадок, чем ответов
на старые вопросы, может быть, уникальные находки более поздних – среднекембрийских
мягкотелых животных скажут нам больше?
Самое богатое местонахождение кембрийских мягкотелых называется Бёрджес
Шейл и находится в Канаде. То, что было там найдено, привело ученых в полное
замешательство. Чего стоят одни названия, которые они дали этим "возмутительным"
тварям, опять, как и вендские существа, не пожелавшим вписываться в красивые
и стройные умозрительные схемы сравнительных анатомов и эволюционистов-теоретиков!
Одно из животных получило название "галлюцигения". Палеонтологи,
видимо, долго протирали глаза и очки, впервые увидев отпечаток этого существа.

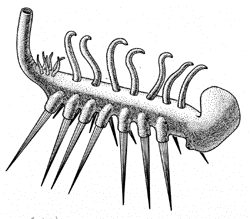
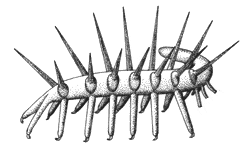
Галлюцигения: отпечаток и две
реконструкции (слева – неправильная, справа – правильная).
На первых найденых отпечатках было видно по три конечности на каждом
сегменте (см. фото): два твердых "шипа" и одно мягкое "щупальце".
Правда, возмутительной вендской "симметрии со сдвигом" здесь уже
не было, симметрия галлюцигении – вполне правильная, билатеральная, но еще
неизвестно, что хуже – ноги в шахматном порядке или по три ноги на сегменте!
Сначала предполагали, что галлюцигения ходила на парных "шипах",
а "щупальца" в один ряд располагались у нее на спине (см. реконструкцию
на рисунке слева).
Потом, к счастью, нашлись другие отпечатки, на которых удалось разглядеть,
что "мягких щупалец" на каждом сегменте было все-таки два, а не
один. Так появилась более правильная (по крайней мере, более правдоподобная)
реконструкция галлюцигении (на рисунке справа). Бедное животное перевернули
вверх тормашками и поставили на парные "щупальца",
а "шипы" стали просто защитными придатками, вроде игл морского
ежа. В таком виде галлюцигения напоминает, хоть и очень отдаленно, современных
онихофор – червеобразных животных, сочетающих признаки членистоногих и кольчатых
червей.
Надо сказать, что в Бёрджес Шейл нашли и других онихофор, больше похожих
на современных.

Айшеайя – онихофора из Бёрджес
Шейл. Голова находится внизу справа; видны приротовые придатки. Возможно,
айшеайя питалась губками, потому что ее отпечатки часто находят вместе с
остатками губок.
Но все "приключения" галлюцигении меркнут по сравнению с уникальной
судьбой другого среднекембрийского монстра – аномалокариса ("аномальной
креветки"). Отдельные части тела этого хищного чудовища (некоторые
экземпляры достигали почти двух метров в длину) сначала находили по отдельности
и описывали как разные организмы. Удивительная "медуза" с дыркой
посередине оказалась ротовым диском аномалокариса. Внутренний край "дырки",
как выяснилось, был усажен острыми зубами. Странные "креветки"
оказались хватательными конечностями необычного хищника. Только когда были
найдены первые полные отпечатки аномалокариса, ученые поняли, как было устроено
это существо (см. рис.)
Реконструкция аномалокариса
Сегментированное тело с "плавательными лопастями" напоминает
некоторых современных кольчатых червей (их конечности-параподии тоже иногда
образуют широкие листовидные лопасти). Однако передние хватательные конечности
(которые сначала были описаны как креветки) и крупные глаза на стебельках
– в точности как у членистоногих.
Мягкотелая фауна Бёрджес Шейл и других подобных местонахождений в Китае,
США и Гренландии открыла перед учеными целую галерею удивительных существ
с разнообразными причудливыми сочетаниями признаков кольчецов и членистоногих.
Все это очень мало похоже на постепенный, плавный переход от одного типа
к другому.
По-видимому, отдельные признаки членистоногих формировались независимо
в разных группах червеобразных организмов. Эти признаки перекомбинировались
и смешивались, образуя самые неожиданные комбинации. Возможно, не обошлось
здесь и без горизонтального межвидового обмена генами (такой перенос могли
осуществлять вирусы).
Большую роль в крупных эволюционных перестройках могут играть мутации
регуляторных генов, которые могут приводить к резким, скачкообразным изменениям
сторения организмов. Например, у современных членистоногих известны мутации,
в результате которых вместо усиков вырастают ноги, брюшные сегменты превращаются
в грудные и так далее. Если в одном из сегментов забарахлит какой-то регуляторный
"ген-переключатель" (причиной может быть единичная "точечная"
мутация регуляторного гена), то это сегмент может приобрести комплекс свойств,
характерных для какого-то другого сегмента. Напримепр, вполне можно допустить,
что в результате подобной мутации от организмов, похожих на аномалокариса
(с единственной парой членистых конечностей, развившихся специально для
схватывания добычи), могли буквально в одночасье произойти формы, имеющие
такие конечности на всех сегментах туловища.
Трудности узнавания переходных форм хорошо видны и на примере знаменитого
"древнекрыла" – археоптерикса, восемь скелетов которого найдено
в Германии в отложениях позднеюрского возраста. Очень долго это существо,
сочетающее признаки хищных динозавров-теропод и птиц, считалось классическим
примером переходной формы и блестящим подтверждением эволюционной теории.
Пока археоптерикс был единственной известной "полуптицей – полурептилией",
особых причин для сомнений у биологов не было: всё казалось простым. От
динозавров произошел археоптерикс, от археоптерикса – настоящие птицы.
Но последовали новые находки, которые, как водится, вместо того чтобы
еще более прояснить ситуацию, безнадежно ее запутали. Во-первых, выяснилось,
что археоптерикс был совсем не одинок. В отложениях мелового периода обнаружилось
множество близких к археоптериксу существ – с такой же смесью птичьих и
динозавровых признаков. Одновременно с ними уже начиная с раннего мела жили
и настоящие птицы – "веерохвостые". В отличие от археоптерикса
и его родичей, у которых хвост был очень длинный, совсем как у рептилий,
и поддерживался скелетом из множества подвижно сочлененных позвонков (поэтому
их называют "ящерохвостыми"), у веерохвостых птиц хвост резко
укорочен, а все хвостовые позвонки слиты в одну небольшую косточку – пигостиль.
Археоптерикс и его родичи – ящерохвостые птицы – вне всяких сомнений были
очень близки к хищным динозаврам, от которых, по всей видимости, и произошли.
Хотя у них выработались многие "птичьи" признаки, никаких переходов
от них к настоящим птицам обнаружить не удалось. У ящерохвостых птиц не
было тенеденции к постепенному приобретению тех птичьих особенностей, которых
им "не хватало", чтобы стать настоящими птицами. Да и в летописи
они появились почти одновременно с веерохвостыми…
Зато обнаружились, причем в большом количестве, разнообразные переходы
от хищных динозавров к ящерохвостым птицам! Оказалось, что многие хищные
динозавры средних и мелких размеров в разное время приобретали различные
птичьи черты. Найдено довольно много видов динозавров с настоящими перьями.
Ученые еще не пришли к согласию, зачем им были нужны эти перья: летать-то
они, скорее всего, не могли. Либо перья возникали как средство планирования
при прыжках с ветки на ветку (у древесных динозавров дромеозаврид), либо
для ускорения бега – у быстро бегающих наземных форм. Недавно найдены совсем
уже ни в какие ворота не лезущие "четырехкрылые" динозавры микрораптор
и криптоволанс, которые, скорее всего, умели неплохо летать. Крупные перья,
явно предназначенные для полета, были у них не только на передних конечностях,
но и на задних! Судя по всему, в разных семействах некрупных хищных динозавров
в юре и раннем мелу шел процесс независимой (параллельной) орнитизации, то есть "оптичивания".
В итоге у бедного археоптерикса не осталось ни одного "птичьего"
признака, который не был бы найден у тех или иных динозавров (раньше такими
признаками считались перья, крючковидные отростки на ребрах и вилочка –
особая V-образная
косточка, возникшая в результате срастания ключиц). Зато те признаки, которые
есть у веерохвостых птиц, но отсутствуют у динозавров, не обнаружены ни
у археоптерикса, ни у его ящерохвостых родственников.
Археоптерикс и его родня, несомненно, произошли от динозавров. Но предками
настоящих птиц они не являются. Это тупиковая ветвь, вымершая в конце мелового
периода и не оставившая потомков.
Предков настоящих (веерохвостых) птиц, очевидно, нужно искать в более
древних эпохах. Известный Российский палеоорнитолог Е.Н.Курочкин убежден,
что птицы произошли вовсе не от пернатых динозавров, не от археоптерикса
и не от подобных ему ящерохвостых птиц, а от каких-то гораздо более древних
пресмыкающихся, скорее всего – от позднетриасовых текодонтов. Эта группа
рептилий была "общим предком" и птиц, и динозавров, и крокодилов.
Лучшим из обнаруженных до сих пор кандидатов на роль птичьего предка среди
текодонтов является протоавис ("первоптица"), найденный в позднетриасовых
отложениях США. Хотя протоавис был наземным животным и не умел летать, у
него имеются важнейшие птичьи признаки, которых нет ни у археоптерикса,
ни у других ящерохвостых, не говоря уже о пернатых динозаврах. Эти признаки
обнаруживаются в особом строении позвонков ("гетероцельные" позвонки),
плечевого и тазового поясов и черепа (увеличенные лобные кости и глазницы).
Остается только надеятся, что "недостающие звенья" – переходные
формы между позднетриасовым протоависом и древнейшими (раннемеловыми) веерохвостыми
птицами – когда-нибудь будут обнаружены!
Иногда "переходные формы" задают ученым загадки совершенно иного
рода. После выхода в свет трудов Дарвина о происхождении человека от обезьяноподобного
предка все ученые, поверившие в эволюцию, страстно хотели найти остатки
"обезьянолюдей". Но найти их удалось не сразу, а хотелось очень
сильно. Настолько сильно, что у кого-то из новоиспеченных эволюционистов,
по-видимому, не выдержали нервы…
В 1909 году в Англии, возле Пильтдауна в графстве Суссекс, были найдены
фрагменты черепа, а в 1912 году – нижняя челюсть долгожданной "переходной
формы". Череп был очень похож на человеческий, но вот челюсть оказалась
очень примитивная, почти такая же, как у современных обезьян.
Находка сразу получила широкую известность и вызвала бурный энтузиазм
ученых во всем мире. Но потом стали появляться другие находки: питекантропы,
синантропы, гейдельбергский человек… Пильтдаунский экземпляр вызывал все
большее недоумение. У других "переходных форм" не было такого
странного сочетания очень примитивной челюсти с почти современной черепной
коробкой. Питекантропы гораздо лучше вписывались в дарвиновскую модель плавного,
постепенного превращения обезьяны в человека, чем пильтдаунский череп…
Спустя 40 лет после сенсационной находки было проведено ее тщательное
переизучение. Вынесенный вердикт нанес серьезный урон престижу всей мировой
науки. Пильтдаунский человек оказался подделкой. Череп современного человека
и челюсть шимпанзе кто-то обработал специальными химическими составами,
чтобы придать им красноватый оттенок, характерный для очень древних костей.
Зубы шимпанзе были аккуратно подточены напильником.
С тех пор антропологи очень тщательно проверяют каждую находку, а противники
теории эволюции – креационисты – до сих пор при каждом удобном случае напоминают
эволюционистам об этом позорном инциденте и заявляют, что и все прочие питекантропы
с австралопитеками, небось, такие же подделки…
Вот как дорого может обойтись один-единственный недобросовестный поступок!
Особенно когда его совершает ученый[2].
[1] от древнееврейских слов "бара" – творить (это второе слово Библии: "Берешит бара Элохим…" – "Вначале сотворил Бог…") и "мин" – "сорт", "разновидность" или "секта" (этим термином – "мин" – называли, например, первых христиан).
[2] Предполагается, что поддельные кости "Пильтдаунского человека" изготовил археолог-любитель Чарлз Доусон, но вина его не является полностью доказанной. К моменту разоблачения он уже давно умер.
Обсудить этот текст можно здесь